Steps in Translation : Initiation
The process of protein synthesis translates the 3-letter alphabet of nucleotide sequences on mRNA into the 20-letter alphabet of amino acids that constitute proteins. The mRNA is translated from its 5′-end to its 3′-end, producing a protein synthesized from its amino (N)-terminal end to its carboxyl (C)-terminal end. Prokaryotic mRNA often have several coding regions (that is, they are polycistronic). Each coding region has its own initiation and termination codon and produces a separate species of polypeptide.
In contrast, each eukaryotic mRNA has only one coding region (that is, it is monocistronic). The process of translation is divided into three separate steps: initiation, elongation, and termination. Eukaryotic translation resembles that of prokaryotes in most aspects. Individual differences are noted in the text.
One important difference is that translation and transcription are temporally linked in prokaryotes, with translation starting before transcription is completed as a consequence of the lack of a nuclear membrane in prokaryotes.
Initiation
Initiation of protein synthesis involves the assembly of the components of the translation system before peptide-bond formation occurs. These components include the two ribosomal subunits, the mRNA to be translated, the aminoacyl-tRNA specified by the first codon in the message, GTP, and initiation factors that facilitate the assembly of this initiation complex (. [Note: In prokaryotes, three initiation factors are known (IF-1, IF-2, and IF-3), whereas in eukaryotes, there are many (designated eIF to indicate eukaryotic origin). Eukaryotes also require ATP for initiation.] The following are two mechanisms by which the ribosome recognizes the nucleotide sequence (AUG) that initiates translation.
1. Shine-Dalgarno sequence: In Escherichia coli (E. coli), a purine-rich sequence of nucleotide bases, known as the Shine-Dalgarno (SD) sequence, is located six to ten bases upstream of the initiating AUG codon on the mRNA molecule (that is, near its 5′-end). The 16S rRNA component of the small (30S) ribosomal subunit has a nucleotide sequence near its 3′-end that is complementary to all or part of the SD sequence. Therefore, the 5′-end of the mRNA and the 3′-end of the 16S rRNA can form complementary base pairs, facilitating the positioning of the 30S subunit on the mRNA in close proximity to the initiating AUG codon (Fig. 1).
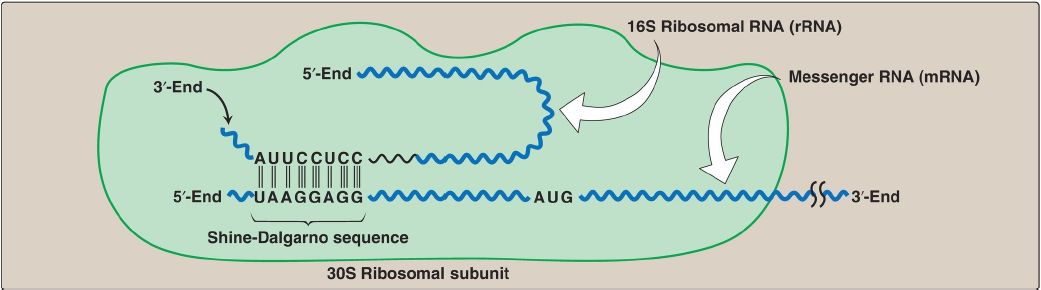
Figure 1: Complementary binding between prokaryotic mRNA Shine-Dalgarno sequence and 16S rRNA. S = Svedberg unit.
2. 5′-Cap: Eukaryotic mRNA do not have SD sequences. In eukaryotes, the small (40S) ribosomal subunit (aided by members of the eIF-4 family of proteins) binds close to the cap structure at the 5′-end of the mRNA and moves 5′→3′ along the mRNA until it encounters the initiator AUG. This scanning process requires ATP. Cap-independent initiation can occur if the 40S subunit binds to an internal ribosome entry site close to the start codon. [Note: Interactions between the cap-binding eIF-4 proteins and the poly-A tail–binding proteins on eukaryotic mRNA mediate circularization of the mRNA and likely prevent the use of incompletely processed mRNA in translation.]
3. Initiation codon: The initiating AUG is recognized by a special initiator tRNA (tRNAi). Recognition is facilitated by IF-2-GTP in prokaryotes and eIF-2-GTP (plus additional eIF) in eukaryotes. The charged tRNAi is the only tRNA recognized by (e)IF-2 and the only tRNA to go directly to the P site on the small subunit. [Note: Base modifications distinguish tRNAi from the tRNA used for internal AUG codons.] In bacteria and mitochondria, tRNAi carries an N-formylated methionine (fMet), as shown in Figure 2. After Met is attached to tRNAi, the formyl group is added by the enzyme transformylase, which uses N10-formyl tetrahydrofolate as the carbon donor. In eukaryotes, tRNAi carries a Met that is not formylated. In both prokaryotic and eukaryotic cells, this N-terminal Met is usually removed before translation is completed. The large ribosomal subunit then joins the complex, and a functional ribosome is formed with the charged tRNAi in the P site. The A site is empty. [Note: Specific (e)IF function as anti-association factors and prevent premature addition of the large subunit.] The GTP on (e)IF-2 gets hydrolyzed to GDP. In eukaryotes, the guanine nucleotide exchange factor eIF-2B facilitates the reactivation of eIF-2-GDP through replacement of GDP by GTP.

Figure 2: Generation of the initiator N-formylmethionyl-transfer RNA (fMettRNAi).
THF = tetrahydrofolate; C = cytosine; A = adenine.





















